July 27, 2015 report
Driving myelination by actin disassembly

(Phys.org)—If a metallurgist wanted to determine how a blade was made they might cut a small cross section, mount, polish, and etch it, and then look at it under a microscope. They could probably tell right away whether it was forged, cast, or perhaps even 3D printed. If the blade was of a particularly high quality, like a Samurai sword, they could see the multiple layers that were created the through laborious cycles of heating, hammering and refolding that are typically used to optimize the grain structure for both toughness, hardness, and grain orientation. The point here is that the structure evident in the Samurai sword cross section didn't just passively polymerize out of thin air like a snowflake, or get made by some other passive thermochemical series of events, it took a more directed driving force.
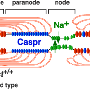
One of the most enduring puzzles in neurobiology is what is the physical force that drives myelin around an axon. When looked at in cross section with an EM microscope, up to 150 or even more tightly wound turns can be seen. At the inner leading edge there is a slightly expanded tip containing a bit of cytoplasm which is called the inner mesaxon or 'tongue'. There is also an outer cytoplasm-containing tongue in compacted myelin sheaths that curiously enough is almost always observed to line up in roughly the same 'clock' position as the inner tongue, but more on that in a bit.
Years of painstaking work have lead to the conclusion that the inner tongue tip is the business end of whatever kind of mechanism it is that is driving myelination. Forgive the generalization, but the current presumption in the field seems to be that if enough details can be learned about the various proteins adorning these tongue domains, and the narrow cytoplasmic incisions and channels connecting them, then the true nature of the mysterious force pushing things forward should pop right out. The idea that the inner tongue is a modified force-generating 'lamellipodium', or growth cone of sorts, has been amply fed by evidence of an actin-based motility mechanism for the initial processes sent out by myelinating glial cells.
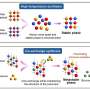
But there's a problem with the idea that this leading edge tongue is just a souped-up actin pseudopod. Recent work by Brad Zuchero and others has shown that while actin assembly powers the initial stage of myelination where axons are first contacted, the actual wrapping stage requires the actin scaffold to be taken down. Their work, just published in Developmental Cell, focuses on the activity of actin-related proteins (Arp2/3) which serve as nucleation sites for new filaments in mouse oligodendrocytes. Although Arp2/3 looks pretty similar to standard-issue monomeric actin, these proteins are actually specialized tinkertoy complexes that bind to existing mother filaments and then seed daughter filaments at precise 70 degree angles.
After demonstrating that Arp2/3-mediated assembly of actin networks is critical in the initial ensheathment process, Brad and the other researchers involved showed Arp2/3 isn't required for the subsequent membrane spreading and wrapping phase. What is required they found, is disassembly of the actin network by actin-severing proteins like gelsolin and cofilin, along with the special services of a ubiquitous protein known as MBP (Myelin basic Protein). Although MBP is known to be essential in kicking off myelination, and may be involved in linking successive layers together, its actual modus operandi is still not clearly known. Based on their recent work, the researchers now suggest that MBP may operate by directly displacing gelsolin and cofilin from the membrane locale where they are typically sequestered in an inactive state.

Okay, since we've just spent a fair amount of space talking about various proteins and aren't really any closer to answering the force question, we should probably return to Samurai swords. It is conceivable that the actual power for myelination, or at least some part of it, comes from outside the glial cell sheath—some force that operates on the sheath like a swordsmith would on metal to fashion its internal geometry. I asked Brad if he thought it might be possible that axons might act as rotating mandrels and spool up myelin from the inside. He concurred that in theory at least, this would be an attractive mechanism because there is ample evidence that axons can and do exert significant torques when they are growing (and perhaps as we have suggested before, also when they are spiking).
However, Brad also quickly countered that this mechanism would not actually be possible. I am heavily paraphrasing him here, but the basic reason is that long long ago the founding fathers had explored the question of the orientation in the myelin wrappings between adjacent nodes going down an axon, and found that it varied randomly. Therefore, a chiral (or handed) axon spooling continuously in one direction could not wind up myelin in different directions in different places. That was a fair enough response, and also one that I had come to expect from the field. Undoubtedly, the first researchers to look at myelin would have been sensible enough to ask the same question and answer it by looking at it in serial sections of a labelled axon.
The peculiar thing is that although I had heard similar responses from other myelination experts, I had never seen a reference—so I pressed Brad for something to attest to his claim. He mentioned that a paper by C. Hildebrand should do the trick. He also recommended I follow up with microscopy expert Mika Simons, who I later realized had also recently published new discoveries in Developmental Cell on actin and myelination. Curiously, Mika pointed me to the same paper with the clarification that 'Fig. 8' should clear everything up. Unfortunately, I found that while the famed Fig. 8 was a scatterplot that purported to show that the number of counterclockwise and clockwise spirals for each 'unit' were equal, the units were not what everybody thought they were. Instead of being adjacent segments down a single axon, a unit was a single glial cell, and the plot was counting all the unit's segments on adjacent axons.
Whether or not this paper is main source of other mythinformation that I have since found on the interwebs (including early sci.neurosci newsgroups from the early nineties stating that myelin orientations were random or 50-50), I can not say. However, I can say that the idea that axon force generating movements—either rotations, or repetitive radial displacements secondary to spikes—can assist glial wrapping is alive and well. One would think that whether or not there still is this seemingly preposterous glial foible lurking at large in the field should be something fairly easy to determine.
In fact, just last year several neuroscience luminaries carried out what is probably the most detailed and exhaustive myelin serial section study to date. Yet somehow they managed to report little more than the seemingly trivial observation that the length of the initial axon segment varied a bit. Maybe these researchers could go back and re-examine things to answer the orientation question once and for all. In covering that particular work we hinted at the idea that not just axons or apical dendrites could be uniquely chiral, but organelles (mitochondria, ER, and centrioles), and consequently also whole cells could be chiral, and furthermore come with different chiralities. Surprisingly, the researcher's own previous work had demonstrated that exact possibility for the helicity in the endoplasmic reticulum.
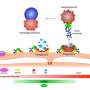
In searching for the mechanism of force generation both Brad and Mika's papers examine various mechanisms of motility in other cell models to see if anything can be learned. For example, in non-adhesive cells like leukocytes, actin-network contractility generates hydrostatic pressure which can induce blebbing protrusions in the membrane. Various mechanisms, like re-hydrating depolymerized actin monomers might be imagined here that could assist in building such pressures. However in oligodendrocytes, not only are the usual suspects, (the motor proteins like the myosin) scarce, actin itself is absent from the compacted myelin area and is only detectable in a thin outer rim of the tongue.
Mika's group proposed and observed that in flat membrane sheets actin polymerization in the tongue could 'inflate' breathing room, and then depolymerize and deflate when the differentiating cell began to extend a myelin webbing across its filopodial outposts. But there is simply no clear mechanism yet for iterating such a cycle through the full turn of a myelin wrap, let alone 150 of them. The have there doubsts about the blebbing model because their precision measurements of membrane stiffness don't jive with the current conception of actin-membrane mechanics during bleb formation.
At an even more fundamental level there is still no good 'theory' of why peripheral nerves get single segment Schwann cells while central nerves get multisegment oligodendrocytes. Compare for example, the fairly recent successful theory of why there are distinct rod and cone photoreceptors that do a similar physical task of hyperpolarizing in response to light. While a good explanation for this may not immediately tell us how they wrap, it should give a much needed sense of proportion to both the physical and molecular events we have described so far.
One pertinent observation here is that when animals are finally ready to myelinate, they can do it pretty fast when needed. It may be the case that it all just comes down to scale—what a given cell can crank out. For example, a Schwann cell sheath that is 15um in diameter x 1500um long with 150 turns would be a volume of 150,000um^3. If both kinds of glial are assumed to operate somewhere near a theoretical metabolic maximum than an oligodendrocyte with 'arms' that are 500um-long x 5um made of 50 turns could put out axon-wrapping units. Complementary studies that explore the staggering complexity of the fuller interspecies glial menagerie operating near their limits may help solve some of the mysteries of actin motility in myelin.
More information: CNS Myelin Wrapping Is Driven by Actin Disassembly, Developmental Cell, www.cell.com/developmental-cel … 1534-5807(15)00400-1
Actin Filament Turnover Drives Leading Edge Growth during Myelin Sheath Formation in the Central Nervous System, Developmental Cell, www.cell.com/developmental-cel … 1534-5807(15)00349-4
Journal information: Developmental Cell
© 2015 Phys.org